History of Biological Psychology
 The history of biological psychology—the study of psychology in terms of bodily mechanisms—is a major part of the history of modern scientific psychology. It has been a prominent field of psychology from the start in Europe and North America and remains a major area of research and instruction in many countries. Over one and one half centuries, biological psychology has found new ways to answer old questions, has tackled important new questions, and has abandoned some problems as poorly defined. Carefully designed behavioral experiments and innovative biomedical techniques have been essential to its progress. Biological psychology has been a site of exchange of concepts. information. and techniques between psychology and the biological sciences.
The history of biological psychology—the study of psychology in terms of bodily mechanisms—is a major part of the history of modern scientific psychology. It has been a prominent field of psychology from the start in Europe and North America and remains a major area of research and instruction in many countries. Over one and one half centuries, biological psychology has found new ways to answer old questions, has tackled important new questions, and has abandoned some problems as poorly defined. Carefully designed behavioral experiments and innovative biomedical techniques have been essential to its progress. Biological psychology has been a site of exchange of concepts. information. and techniques between psychology and the biological sciences.
The current scope of biological psychology is broad including the following topics:
- Evolution of brain and behavior;
- Development of the nervous system and behavior over the life span;
- Psychopharmacology;
- Sensory and perceptual processes;
- Control and coordination of movements and actions;
- Control of behavioral states (motivation), including sex and reproductive behavior, and regulation of internal states;
- Biological rhythms and sleep;
- Emotions and mental disorders;
- Neural mechanisms of learning and memory, language and cognition;
- Recovery of function after damage to the nervous system.
Developing from biological psychology and overlapping with parts of it are such fields as behavior genetics and hormones and behavior.
Origins of Biological Psychology
During much of its history. biological psychology was known as “physiological psychology.” but we should note two meanings of this term. First. physiological psychology was the name Wilhelm Wundt and his contemporaries used for the new. scientific psychology. The preface to Wundt’s most important book. Grundzüge der physiologischen Psychologie (Principles of Physiological Psychology. 1873-1874). stated his purpose:
The work which I here present to the public is an attempt to mark out a new domain of science…. The question may be raised. whether the time is yet ripe for such an undertaking. The new discipline rests upon anatomical and physiological foundations which. in certain respects, are themselves very far from solid; while the experimental treatment of psychological problems … [is] still at its first beginnings. (1948. p. 248)
Wundt’s book treated general experimental psychology. which he hoped to demonstrate could be as scientific as physiology. Physiology itself was one of the streams of investigation and theory that joined to form the new psychology. Other lines that led to psychology included human and comparative neuroanatomy, study of sensation, and sociomedical studies.
By the 1920s, physiological psychology was acquiring its second, more restricted meaning: the study of behavior in terms of its biological mechanisms. Textbooks on physiological psychology in this sense began to appear in the 1930s. The discipline of physiology also became more restricted during the twentieth century as new sciences split from it—sciences such as endocrinology, genetics, and biochemistry. Thus the term biological psychology arose as being more appropriate to designate the relations of psychology to the life sciences. Textbooks entitled Biological Psychology began to appear in the 1980s. Other terms sometimes used as alternatives to biological psychology or “biopsychology” include physiology of behavior and behavioral neuroscience. Currently. biological psychology encompasses four main aspects: biological mechanisms of behavior; the development of behavior and its biological mechanisms over the life span; the evolution of behavior and its biological mechanisms; and applications. which entail both tests of the adequacy of formulations and use of findings to benefit health and quality of life (Rosenzweig. Leiman. & Breedlove, 1999).
Persistent Themes in Biological Psychology
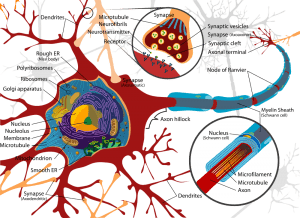 Biological psychology continues some of the main lines of theorizing and research that led to the development of psychology as an independent discipline. These lines include the explanation and interpretation of behavior in terms of the physiology of the nervous system; the development of psychology on the basis of evolution; the localization of function in the nervous system; and the plasticity of the nervous system.
Biological psychology continues some of the main lines of theorizing and research that led to the development of psychology as an independent discipline. These lines include the explanation and interpretation of behavior in terms of the physiology of the nervous system; the development of psychology on the basis of evolution; the localization of function in the nervous system; and the plasticity of the nervous system.
Physiological Explanations of Behavior
Rene Descartes (1596-1650) was an important forerunner of modern psychology who sought to locate mental functions in the nervous system and to explain aspects of behavior in terms of physiological mechanisms. Another pioneer was the physician David Hartley (17051757). a philosopher by avocation. In his Observations on Man (1749), Hartley attempted to integrate the current facts and theories of anatomy and physiology with concepts of associationistic philosophy. Apparently the irst to use the term physiological psychology in a book title was English physician Robert Dunn (1858). The philosopher and psychologist Alexander Bain (18181903) sought to establish solid connections between psychology and physiological knowledge in his textbooks of the 1850s, The Senses and the Intellect and The Emotions and the Will. Combined and abridged as Mental and Moral Science (1868), this was the major textbook of psychology in Great Britain for almost 50 years. Among other anticipations of later work. Bain suggested that memory formation involves growth of what we now call synaptic junctions:
for every act of memory. every exercise of bodily aptitude, every habit, recollection. train of ideas. there is a specific grouping or coordination of sensations and movements, by virtue of specific growths in the cell junctions. (1872, p. 91)
Evolution
Herbert Spencer espoused the concept that evolution of mind is central to psychology in his influential book. Principles of Psychology (1855). This was shortly before Charles Darwin and Alfred Russel Wallace formally proposed the theory of evolution by natural selection. Darwin also envisaged the importance of evolution for psychology. In On the Origin of Species, Darwin. who had not yet read Spencer’s Principles of Psychology, looked forward to the time when “Psychology will be based on a new foundation, that of the necessary acquirement of each mental power and capacity by gradation” (1859, p. 113). Darwin contributed directly to psychology in later writings: The Descent of Man (1871), The Expression of Emotions in Man and Animals (1872), and the first detailed case study of child development, ‘A Biographical Sketch of an Infant” (1877). He has been credited with “overwhelming importance in the development of psychology as a branch of evolutionary biology in the three separable areas of comparative psychology, functional psychology, and the study of the nervous system” (Young, 1970, p. 191). Once the laws of heredity were recognized at the beginning of the twentieth century and the mechanisms of heredity were worked out later in the century, evolution of aspects of behavior became a fruitful subject of investigation, and behavior genetics became a productive field.
Some vestiges of older thinking persisted into the twentieth century, however, such as the Aristotelian concept that animals can be arranged in a scala naturae, a single ladder-like progression from the simplest to the most complex and perfected, with humans at the pinnacle. Thinkers in the nineteenth century referred to this as the phylogenetic scale, and this concept was still reflected in Clifford Morgan’s influential textbook, Physiological Psychology (1943). In the twentieth century, evolutionary biologists developed the concept of a widely branching phylogenetic tree that replaced the phylogenetic scale. Current textbooks in biological psychology and related ields refrain from suggesting that evolution necessarily means an increase in complexity and avoid such terms as primitive species or advanced species (Butler & Hodos, 1996; Rosenzweig et al., 1999).
Localization of Function
A critical advance was the discovery of the difference between sensory and motor nerves, made independently by Charles Bell in 1811 and Francois Magendie in 1822. Bell and Magendie showed that cutting dorsal spinal roots made a limb insensitive but did not prevent motion, whereas cutting the ventral roots made the limb immobile. Previously, many physiologists had believed that spinal nerves carried sensory and motor messages in both directions at once. Bell also argued for further subdivision among the nerves, claiming that the five senses are mediated by different nerves, thus anticipating Johannes Müller ‘s 1826 doctrine of “specific energies of nerves” of the different senses. This doctrine was later extended by Müller ‘s student Hermann Helmholtz and others to account for differences within a sensory modality (e.g., hues, tones, and different skin sensations such as touch, warmth, and cold).
The “law of spinal roots” can be considered a special case of the more general idea of localization of function in the nervous system that gained much attention in scientific circles early in the nineteenth century. The first formal proponent of the concept of localization was the physician and anatomist Franz Joseph Gall, who called his system “organology” because he believed the brain to be composed of a number of different organs with different functions (1810-1819). Johann Spurzheim, who was associated for several years with Gall, popularized the term “phrenology,” which Gall refused to use. Gall believed he could identify and localize 27 faculties or abilities in different parts of the human cerebral cortex. On the basis of comparative studies of other mammals, he believed he could demonstrate and localize nineteen of these faculties in animals. Both Gall and Spurzheim attempted to locate the organs of the brain by studying the protuberances in the skulls of people who had particular talents or striking characteristics or by assessing individuals with unusually shaped skulls.
A prominent anatomist-physiologist, Marie Jean Pierre Flourens, attacked the claims of Gall and Spurzheim. Making experimental lesions in the brains of a variety of species, especially birds, and stimulating parts of the brain, he found only limited evidence for localization of function (Flourens, 1824). Flourens found that each sense organ has its own representation in the cerebrum, and he also noted that, following localized brain lesions, animals would lose certain functions but could recover them completely. He therefore concluded that the cerebral cortex functions as a whole and that all of its parts are responsible for intelligence, the will, and perception. As one neurohistorian notes: “it is possible to think of Gall as the visionary who had the right idea but the wrong method, and of Flourens as the laboratory scientist with the better method but the wrong theory” (Finger, 1994, p. 36).
Paul Broca’s evidence for localization of speech function in the left frontal cortex (1865) gained wide acceptance for the concept of localization of function in the cortex and initiated a long period of fruitful studies. Further developments, associated with names such as Eduard Hitzig and David Ferrier, led to a concept of the brain as being composed of fixed compartments. Thus anatomist Korbidian Brodmann (1909) divided the human cerebral cortex into 52 regions based on cytoarchitectonic criteria and suggested that each region has a special function. Further anatomical research indicated even finer subdivisions, with some investigators claiming as many as 200 separate cortical fields. Among the skeptics about functional mapping and diagramming was Karl Lashley, who showed that considerable variation existed among the primary visual cortices of monkeys (Lashley & Clark, 1946).
Research by Roger Sperry, a student of Lashley’s. on development of brain connections in amphibians was understood by many neurobiologists to mean that connections in the brain are formed independently of activity or experience and are programmed by a set of recognition molecules on each neuron (Sperry. 1963). Sperry. however. limited inflexibility to the basic neural projection systems and believed that learning could alter connections in higher brain regions (Sperry. 1951).
Much current research on localzation of function in the human brain utilizes methods of noninvasive brain imaging. Brain imaging. done either when a person is at rest or performing a mental task, shows activity over much of the brain. It is only when brain activity recorded under a “baseline” condition is subtracted from activity during the task that certain brain regions are seen to have heightened or depressed levels of activity. Psychologists contribute to this research in part by designing specific tasks to study particular functions.
Plasticity of the Nervous System
Early in the nineteenth century theorists differed about whether plasticity was a property of the nervous system. Thus Gall emphasized the innate development of the different “organs” of the cortex, each of which he hypothesized to correspond to a different mental faculty. Gall rejected Spurzheim’s idea that humankind is almost indeinitely perfectible and that exercise or education could influence the development of the faculties or the organs of the brain (Gall, vol. 4, 1819, pp. 252-256). Jean-Baptiste Lamarck. the originator of the ill-fated doctrine of evolution through inheritance of acquired characteristics, held that the brain and each of its special regions develops through appropriate use of the related faculties. and he criticized Gall’s belief that brain development is determined innately (1809/1914).
Hermann Ebbinghaus’s (1885) demonstration that learning and memory can be measured encouraged psychologists and neurobiologists to speculate further about neural mechanisms of learning and memory. Thus William James (1890) speculated that learning is related to anatomical changes at neural junctions, as did neurobiologists Eugenio Tanzi (1893), Santiago Ramon y Cajal (Cajal, 1894), and Charles Sherrington (Foster & Sherrington. 1897). In the same publication, Sherrington proposed the term “synapses” for these junctions (Foster & Sherrington, 1897, p. 929).
Initial publications about learning in animals by Edward Thorndike (1898) and Ivan P. Pavlov (1906) encouraged research on neural mechanisms of learning. Psychologist Shepard I. Franz was the first to seek to determine the site of learning in the brain by combining Thorndike’s methods of training and testing animals with the technique of inducing localized brain lesions. Franz later inducted Karl Lashley, and through Lashley many others, into research on this topic. Franz also worked on rehabilitation of brain-injured soldiers, claiming success in obtaining functional recovery even in cases of long-standing paralysis: “we should probably not speak of permanent paralysis, or of residual paralysis, but of uncared for paralysis” (Franz, Sheetz, & Wilson. 1915).
By the mid-twentieth century, the study of brain plasticity and neural mechanisms of learning and memory seemed to have stalled. Lashley published a pessimistic review in 1950. Hans-Lukas Teuber stated that
…the absence of any convincing physiological correlate of learning is the greatest gap in physiological psychology. Apparently, the best we can do with learning is to prevent it from occurring, by intercurrent stimulation through implanted electrodes … , by cerebral ablation … , or by depriving otherwise intact organisms, early in life, of normal sensory influx. (1955, p. 267)
In fact, around the middle of the twentieth century, major advances were beginning to occur in research on the neural mechanisms of learning and memory. Some of these resulted from recently developed techniques, such as single-cell electrophysiological recording, electron microscopy, and use of new neurochemical methods. Another major influence encouraging research on neural mechanisms of learning and memory was Donald O. Hebb’s 1949 monograph, The Organization of Behavior. Hebb was more optimistic about possible synaptic changes in learning than Lashley was. Hebb noted some indirect evidence for neural changes and did not let the absence of conclusive evidence deter him from reviving hypotheses about the conditions that could lead to formation of new synaptic junctions and underlie memory. In essence, Hebb’s hypothesis of synaptic change underlying learning resembled James’s formulation: “When two elementary brain-processes have been active together or in immediate succession, one of them, on recurring, tends to propagate its excitement into the other” (James, 1890, p. 566). Hebb’s “dual trace hypothesis” also resembles the “consolidation-perseveration” hypothesis formulated in 1900 by Georg Muller and Alois Pilzecker. Much current neuroscience research concerns properties of what are now known as Hebbian synapses.
In the early 1960s two experimental programs announced findings demonstrating that the brain can be altered measurably by training or differential experience. First was the demonstration by an interdisciplinary group at Berkeley that both formal training and informal experience in varied environments led to measurable changes in neurochemistry and neuroanatomy of the rodent brain (Krech, Rosenzweig, & Bennett, 1960; Rosenzweig, Krech, & Bennett, 1961; Rosenzweig, Krech, Bennett, & Diamond, 1962). Soon after came the report of David Hubel and Torsten Wiesel that occluding one eye of a kitten led to reduction in the number of cortical cells responding to that eye (Hubel & Wiesel, 1965; Wiesel & Hubel, 1963, 1965). Later reports of the Berkeley group and others demonstrated specific changes in anatomy of neurons and synapses as a result of training or enriched experience. and such changes were induced in adults as well as in young animals.
In spite of initial skepticism. by the early 1970s some neurobiologists began to accept the reports that significant changes in the brain can be caused by training or exposure of animals to differential experience. Other neurobiologists continued into the 1980s to believe that neural connections in the adult brain remained fixed. The report of Wiesel and Hubel (1965) that changes can be induced in the visual system only during a critical period early in life served to solidify the belief of many neurobiologists that neural connections in the adult brain are fixed and do not vary as a result of training. Later. however. investigators found that modifying sensory experience in adult animals could alter both receptive fields of cells and cortical maps (see reviews by Kaas. 1991; Weinberger. 1995).
In the current synthesis. localization of function and brain plasticity are not incompatible. For example. knowing the modal location of somatosensory representation in the cortex helps investigators to study in detail how the “cortical map” in that region changes as a result of specific training of the fingers.
Training, Professional Organizations, and Journals
 Professional positions in biological psychology are mainly in academic and research institutions. Training for most of these positions requires a doctorate. and many departments of psychology that offer doctorates have programs in biological psychology or behavioral or cognitive neuroscience. The number of doctoral programs in physiological and/or biological psychology in the United States and Canada almost doubled from 1973 to 1992. The National Research Council lists more than 100 research doctorate programs in neuroscience in the United States. Some of these programs are located in departments of psychology. In fact. psychology departments were mentioned as sites of neuroscience programs more frequently than any other specific departments. A plurality of the neuroscience programs were listed as interdisciplinary or multidisciplinary. and many of these mentioned psychology as one of the disciplines involved. Overall. it appears that psychology is participating in the growth of neuroscience rather than being overshadowed by it. (See Rosenzweig, 1998, for a discussion of relations between psychology and neuroscience.)
Professional positions in biological psychology are mainly in academic and research institutions. Training for most of these positions requires a doctorate. and many departments of psychology that offer doctorates have programs in biological psychology or behavioral or cognitive neuroscience. The number of doctoral programs in physiological and/or biological psychology in the United States and Canada almost doubled from 1973 to 1992. The National Research Council lists more than 100 research doctorate programs in neuroscience in the United States. Some of these programs are located in departments of psychology. In fact. psychology departments were mentioned as sites of neuroscience programs more frequently than any other specific departments. A plurality of the neuroscience programs were listed as interdisciplinary or multidisciplinary. and many of these mentioned psychology as one of the disciplines involved. Overall. it appears that psychology is participating in the growth of neuroscience rather than being overshadowed by it. (See Rosenzweig, 1998, for a discussion of relations between psychology and neuroscience.)
Among the scholarly and professional organizations and journals related to biological psychology are the following: Division 6 of the American Psychological Association (APA). founded in 1945. when the APA reorganized. as the Division of Physiological and Comparative Psychology and renamed the Division of Behavioral Neuroscience and Comparative Psychology in 1995. The APA publishes the journal Behavioral Neuroscience, known from 1921-1982 as the Journal of Comparative and Physiological Psychology. Other organizations and publications in which biological psychologists participate include the Society for Neuroscience and its Journal of Neuroscience. the European Brain and Behaviour Society and its journal Behavioural Brain Research. the Forum of European Neuroscience Societies and its European Journal of Neuroscience. and the International Brain Research Organization (IBRO).
There are several current textbooks in the field. Reviews of research in biological psychology appear regularly in Annual Review of Neuroscience. Annual Review of Psychology, and Trends in Neurosciences.
Bibliography:
- A. (1868). Mental and moral science. London: Longmans.
- A. (1872). Mind and body: The theories of their relation. London: King.
- P. (1865). Sur le siege de la faculte du langage articuie [On the site of the faculty of speech]. Bulletin de la Societe d’Anthropologie, 6, 337-393.
- K. (1909). Vergleichende Lokalisationslehre der Grosshirnrinde in ihren Prinzipien dargestellt auf Grund des Zellenbaues [Comparative study of localization in the cerebral cortex in terms of principles based on cytoar-chitectonics]. Leipzig, Germany: Barth
- A. B.. & Hodos. W. (1996). Comparative vertebrate neuroanatomy: Evolution and adaptation. New York: Wiley-Wiss.
- Cajal, R. S. (1894). La fine structure des centres nerveux [The microscopic structure of the central nervous system]. Proceedings of the Royal Society, London. 55, 444468.
- C. (1859). On the origin of the species by means of natural selection. or the preservation of favoured races in the struggle for life. London: Murray.
- C (1871). The descent of man and selection in relation to sex. London: Murray.
- C. (1872). The expression of the emotions in man and animals. London: Murray.
- C. (1877). A biographical sketch of an infant. Mind: Quarterly Review of Psychology and Philosophy, 2, 285-294.
- Dennis, W. (Ed.) (1948). Readings in the history of psychology. New York: Appleton-Century-Crofts.
- R. (1858). An essay on physiological psychology. London: Churchill.
- Ebbinghaus, H, (1885). Ueber das Gedachtnis [On memory]. Leipzig, Germany: Dunker & Humbolt.
- Finger, S. (1994). Origins of neuroscience: A history of explorations into brain function. New York: Oxford University Press.
- Flourens, P. (1824).Recherches experimentales sur les pro-prietes et les fonctions du systeme nerveux dans les animaux vertebras [Experimental research on the properties and the functions of the nervous system in vertebrate animals]. Paris: Balliere.
- Foster, M .. & Sherrington, C. S. (1897). Part III. The central nervous system: A text-book of physiology (pp. 915-1252). London: Macmillan.
- Franz, S. I.. Sheetz, M. E., & Wilson, A. A. (1915). The possibility of recovery of motor functions in long-standing hemiplegia. Journal of the American Medical Association, 65, 2150-2154.
- Gall, J. F. (1810-1819). Anatomie et physiologie du systeme nerveux en general. et du cerveau en particulier. avec des observations sur la possibilite de reconnoitre plusieurs dispositions intellectuelles et morales de l’homme et des animaux par la configuration de leurs tetes [The anatomy and physiology of the nervous system in general. and of the brain in particular. with some observations on the possibility of recognizing several intellectual and moral dispositions of man and animals by the configuration of their heads]. (Vols. 1-4). Paris: Maze.
- Hartley, D. (1749). Observations on man. his frame. his duty. and his expectations. London: Leake & Frederick.
- Hebb, D. O. (1949). The organization of behavior: A neuropsychological theory. New York: Wiley.
- HubeL D. H.. & Wiesel. T. N. (1965). Binocular interaction in striate cortex of kittens reared with artificial squint. Journal of Neurophysiology. 28, 1041-1059
- James, W. (1890). Principles of psychology. New York: Holt.
- J. H. (1991). Plasticity of sensory and motor maps in adult animals. Annual Review of Neuroscience, 14, 137-167.
- Krech, D., Rosenzweig, M. R., & Bennett. E. L. (1960). Effects of environmental complexity and training on brain chemistry. Journal of Comparative and Physiological Psychology, 53, 509-519.
- Lamarck, J. B. (1914). Zoological philosophy. (H. Elliott, Trans.). London: Macmillan. (Original work published 1809)
- K. S. (1950). In search of the engram. Symposia of the Society for Experimental Biology, 4, 454-482.
- Lashley, K. S. & Clark, (1946). The cytoarchitecture of the cerebral cortex of Ateles: A critical examination of architectonic studies. Journal of comparative neurology, 85, 223-305.
- Morgan, C. T. (1943). Physiological psychology. New York: McGraw-Hill.
- Pavlov, I. P. (1906). The scientific investigation of the psychical faculties or processes in the higher animals. 24, 613-619. The same article also appeared in Lancet, 2. 911-915.
- M. R. (1998). Reciprocal relations between psychology and neuroscience. In M. Sabourin, F. Craik, & M. Robert (Eds.), Proceedings of the Twenty-sixth International Congress of Psychology: Vol. 2. Advances in psychological science: Biological and cognitive aspects (pp. 215-236). Hove, England: Psychology Press.
- M. R., Krech. D. . & Bennett, E. L. (1961). Heredity, environment, brain biochemistry, and learning. In Current trends in psychological theory (pp. 87-no). Pittsburgh, PA: University of Pittsburgh Press.
- Rosenzweig, M. R., Krech, D., Bennett, E. L., & Diamond, M. C. (1962). Effects of environmental complexity and training on brain chemistry and anatomy: A replication and extension. Journal of Comparative and Physiological Psychology, 55, 429-437.
- Rosenzweig, M. R.. Leiman, A. L., & Breedlove, S. M. (1999). Biological psychology: An introduction to behavioral, cognitive, and clinical neuroscience (2nd ed.). Sunderland. MA: Sinauer.
- Spencer, H. (1855). The principles of psychology. London: Longmans.
- Sperry, R. W. (1951). Mechanisms of neural maturation. In S. S. Stevens (Ed.), Handbook of experimental psychology (pp. 236-280). New York: Wiley.
- Sperry, R. W. (1963). Chemoaffinity in the orderly growth of nerve fiber patterns and connections. Proceedings of the National Academy of Sciences USA. 50, 703-710.
- Tanzi, E. (1893). I fatti e le induzioni nell’odierna istologia del sistema nervoso [Facts and conclusions from present-day history of the nervous system]. Revista Sperimentale di Freniatria e di Medicina Legale, 19, 419472.
- Teuber, H.-L. (1955). Physiological psychology. Annual Review of Psychology, 6, 267-296.
- Thorndike, E. L. (1898). Animal intelligence: An experimental study of the associative processes in animals. Psychological Monographs. 8, 1-109.
- Weinberger, N. M. (1995). Dynamic regulation of receptive fields and maps in the adult sensory cortex. Annual Review of Neuroscience. 18, 129-158.
- T. N., & Hubel, D. H. (1963). Single-cell responses in striate cortex of kittens deprived of vision in one eye. Journal of Neurophysiology. 26. 1003-1017.
- T. N.. & Hubel, D. H. (1965). Comparison of the effects of unilateral and bilateral eye closure on cortical unit responses in kittens. Journal of Neurophysiology, 28. 1029-1040.
- W. (1873-1874). Grundziige der physiologischen Psychologie [Principles of physiological psychology]. Leipzig, Germany: Engelmann. [Part of the author’s preface, translated by Edward B. Titchener, appears on p. 248 in W. Dennis (Ed.) (1948). Readings in the history of psychology. New York: Appleton-Century-Crofts.]
- Young, R. M. (1970). Mind, brain. and adaptation in the nineteenth century. New York: Oxford University Press.